Article Text

Abstract
Introduction: Interaction between genetic background and oxidative environmental stimuli in the pathogenesis of human lung disease has been largely unexplored.
Methods: A prospective epidemiological study was undertaken in 253 coal miners. Intermediate quantitative phenotypes of response to oxidant exposure, including erythrocyte glutathione peroxidase (GSH-Px) and catalase activities, were studied. Oxidant exposures studied were smoking habits and cumulative dust exposure assessed by job history and ambient measures. Disease phenotypes included subclinical computed tomography score at the first survey and x ray profusion grades twice, five years apart, to assess established coal workers’ pneumoconiosis (CWP). Miners were genotyped for common functional polymorphisms in the gene for tumour necrosis factor α (TNF) and lymphotoxin α (LTA), two proinflammatory cytokines that have been implicated in the pathogenesis of chronic lung diseases.
Results: Regarding gene-environment interaction on intermediate phenotypes, results showed interaction of a promoter polymorphism at the –308 position in TNF with occupational exposure on erythrocyte GSH-Px activity with a significant association in those with high exposure (p=0.003), whereas no association was observed among those with low exposure (interaction p=0.06). Regarding gene intermediate phenotype interaction on clinical outcome, results showed an association of CWP prevalence with an NcoI polymorphism in LTA in those with low catalase activity (p=0.05), whereas no association was observed in those with high activity (interaction p=0.03). No other significant association was observed.
Conclusion: The results suggest that interactions of genetic background with environmental exposure and intermediate response phenotypes are important components in the pathogenesis of CWP.
- TNF
- LTA
- oxidative stress
- pneumoconiosis
Statistics from Altmetric.com
Genetic factors that modify the response to oxidative environmental stimuli in humans are largely unknown. The overproduction of reactive oxygen species (ROS) owing to major environmental factors such as cigarette smoking and chronic exposure to particles induces chronic airway inflammation.1–3 Inflammation is fundamental to the pathogenesis of many chronic lung diseases including asthma, chronic obstructive pulmonary disease (COPD), and pneumoconiosis/fibrosis.4,5 Coal workers’ pneumoconiosis (CWP) is an inflammatory lung disease caused by chronic inhalation of coal dust, and ROS are thought to be important in disease pathogenesis. The source of ROS is proposed to derive directly from the coal dust, and indirectly from activated inflammatory cells such as alveolar macrophages and polymorphonuclear leucocytes.6
The specific mechanisms of oxidative injury are becoming increasingly clear. ROS and specific proinflammatory cytokines such as tumour necrosis factor alpha (TNF-α) activate transcription factors including nuclear factor-κB (NF-κB) via specific intracellular signalling pathways. Translocation of transcription factors to the nucleus from the cytoplasm subsequently initiates transcription in genes with multiple inflammatory functions, including cytokines (TNF, IL-4, IL10, etc), and antioxidant enzymes such as glutathione peroxidase, superoxide dismutases, and catalase.7,8
Interestingly, only some people similarly exposed to environmental oxidative stimuli (for example, cigarette smoke and particles) develop lung disease, and the determinants of susceptibility are poorly understood. Intrinsic factors such as genetic background may be important determinants of interpersonal susceptibility to disease pathogenesis. A number of candidate genes for susceptibility to chronic pulmonary diseases have been investigated. The –308 G/A SNP in the promoter region of the gene (TNF) for TNF-α has been studied frequently. TNF is located in tandem with the lymphotoxin-α gene (LTA) in the class III region of the 3.6 Mb human major histocompatibility complex (MHC) on chromosome 6p21.31,9 and the TNF gene superfamily plays an important role in inflammation. The –308 promoter polymorphism has been associated with subphenotypes of asthma, COPD, fibrosis, and CWP in some studies,10–14 but not in others.15,16 Other genetic factors of CWP have also been studied.17 Similarly, a functional NcoI RFLP polymorphism in LTA was associated with sarcoidosis in the study of Yamaguchi et al,18 but it has not been replicated. Discordant results may be related to differences in population characteristics. Differential effects of the same environmental exposure on subjects with different genotypes may change the magnitude or the direction of the associations but, unfortunately, none of the studies mentioned above has considered the effects of gene-environment interactions in the disease aetiology or included biological markers of the response to oxidative stress.
The overall hypothesis of the present study was that TNF −308 and LTA NcoI polymorphisms modify the pulmonary responses to oxidants in coal miners differently exposed to cigarette smoke and coal mine dust. To test this hypothesis, genotype-environment interactions on intermediate quantitative phenotypes of response to oxidative stress were specifically investigated. Further, genotype intermediate phenotype interactions on disease phenotypes were studied. The response to oxidative stimuli (coal dust, cigarette smoke) was assessed by biological intermediate quantitative phenotypes (enzyme activities), and detailed information on coal workers’ pneumoconiosis (assessed from computed tomography and twice by chest radiography five years apart). A major strength of this study is the quantitative assessment of environmental exposure to oxidative stimuli that permits evaluation of gene-environment interaction.
SUBJECTS AND METHODS
Subjects studied took part in a longitudinal epidemiological study, including quantitative environmental measurements, biological assessment of response to oxidative stress, and radiological assessment of coal workers’ pneumoconiosis (CWP), including subclinical phenotypes.
Study sample
The population studied consisted of 259 unrelated coal miners recruited through a standardised protocol19 in a French coal mine (Houilléres du Bassin de Lorraine, in north eastern France) based on a selection contrasted by exposure and disease status. The sample included 240 active coal miners aged 34-50 years in 1990, including 80 subjects heavily exposed to underground coal dust (≥10 years at the coal face) with chest x ray classified 0/1 or 1/0; 80 subjects exposed to underground coal dust with normal chest x ray classified 0/0; and 80 subjects slightly exposed with normal chest x ray. Also included were 19 retired miners with CWP, that is, chest x ray classified 1/1 or over in 1994 and an evolution of two stages of x ray grade within the past 10 years. The active coal miners were examined three times (1990, 1994, and 1999) and the retired miners twice (1994 and 1999). Most of the analyses were performed on the active miners using 1994 and 1999 data. The retired miners with established disease have been included for detailed analysis in relation to disease. Out of the 240 active miners recruited in 1990, 234 took part in the 1994 survey and 227 in 1999. Out of the 19 retired miners recruited in 1994, 18 were re-examined in 1999. Based on available data, comparison of those lost to follow up and the remainder did not show any differences regarding exposure, genotype, or disease.
Environment
Low or high current exposure based on job description and cumulative exposure assessed through job history and dust measurements at various sites of the mine were recorded. High current exposure refers to miners working at the coal face, mining, stope, or drift advance, and low exposure to those working at ventilation maintenance, pumping, haulage, shaft, stock equipment, or safety. Cumulative personal exposure to dust was estimated from each person’s job history and from dust measurements at various sites of the coal mine according to the calculation previously described.20 The estimates were summations of each dust measurement (mg/m3) for the respective time spent in each job. Estimated cumulative exposures to dust were calculated until 1999 and expressed as mg/m3 × year. Most of the analysis is based on high and low cumulative exposure using the same threshold (>71 mg/m3 × year) as previously.21 Detailed smoking habits were recorded by standardised questionnaire.
Radiographic examination
Chest x rays and computed tomography (CT) scans were performed to evaluate disease status. International Labour Office (ILO) classification in 12 profusion grades (0/-, 0/0, 0/1, 1/0, 1/1, 1/2, 2/1, 2/2, 3/1, 3/2, 3/3, 3/+) was used at the three examinations for all the miners.22 Pneumoconiosis is defined by a grade ≥1/1. Five categories were used in analyses: 0/- or 0/0, 0/1, 1/0, 1/1 or 1/2, and 2/1 or over. No subject was grade 3/1, 3/2, or 3/3 and only one subject was grade 3/+. CT scans were performed twice for all active miners at recruitment, that is, 240 according to standardised methods.23,24 CT scans were interpreted by two experienced radiologists blinded to individual exposure and x ray findings. Analyses were based on the profusion score already described25 computed as the average over six lung zones (upper, middle, lower parts of both lungs) of profusion score rated from 0 to 3 (absent, rare, intermediate, high profusion). The study was approved by the appropriate ethical committee and written consent was obtained from all subjects.
Assessment of response to oxidative stimuli
Total plasma antioxidant concentration (ascorbate, protein thiols, bilirubin, urate, and α tocopherol), plasma selenium (Se) concentration, Cu++/Zn++ superoxide dismutase (Cu++/Zn++ SOD), glutathione peroxidase (GSH-Px), and catalase activities were determined at the 1994 survey by methods previously described.21,26 Activities were measured in the two weeks following storage and expressed as U/g Hb (Cu++/Zn++ SOD, GSH-Px) or k/g Hb (catalase) in erythrocytes, and U/l (GSH-Px) in plasma. All samples were analysed in duplicate or triplicate and the precision (coefficient of variation) was <10%. Measures were available for 227 miners who did not differ regarding exposure or disease from those without.
Genotyping
Genomic DNA was isolated from 300 μl aliquots of stored blood using the Wizard Genomic DNA Purification Kit from Promega Corporation (Madison, WI). The manufacturer’s recommended protocol was modified slightly as follows: (1) centrifugation was performed at 14 000 × g; (2) incubation and centrifugation periods were doubled; (3) rehydration in TE was achieved with gentle rotation at room temperature for one week. Amplification for both polymorphisms was performed using our previously optimised conditions, which include 60 ng DNA, 1.5 mmol/l MgCl2, 0.4 μmol/l forward and reverse primers, 200 μmol/l dNTPs, and 0.5 U Taq polymerase in a 25 μl reaction volume.
For determination of the A→G polymorphism at position –308 within TNF, we synthesised primers according to Zhu et al.27 Briefly, the forward primer for amplification was designed with a mismatch that inserts a StyI restriction site into the wild type allele. Thirty-five cycles of amplification with an annealing temperature of 59°C were performed. PCR products were subsequently digested with StyI at 37°C for two hours. Fragments were resolved on 3% NuSieve/TBE gels stained with ethidium bromide. The resultant product sizes were 20 and 123 bp for wild type alleles (allele 1), 143 bp for mutant alleles (allele 2), and 20, 123, and 143 bp for heterozygote alleles.
The NcoI RFLP within the first intron of LTA was analysed using the primer design and temperature parameters of Stuber et al.28 Briefly, 37 cycles of 95°C denaturation for 30 seconds, 68°C annealing for 30 seconds, and 74°C extension for 42 seconds were performed. PCR products were digested at 37°C for one hour with NcoI. Fragments were resolved on 2% agarose gels stained with ethidium bromide. The resultant product sizes were 196 and 586 bp for LTA mutant alleles (allele 1), 782 bp for LTA wild type alleles (allele 2), and 196, 586, and 782 bp for heterozygote alleles. Genotypes were determined for 231 miners. Comparisons of genotyped miners with those not (n = 22) did not show differences regarding exposure and disease.
Statistical methods
Standard statistical tests were used including χ2 (or Fisher exact test when appropriate), analysis of variance, and multiple regression analysis. The interaction between genetic polymorphisms with exposure to oxidative stress and health indicators was statistically tested. Significance was assessed at the 5% two sided level. All analyses were performed with the SAS statistical software.29
RESULTS
Genotype distributions and environmental exposures to oxidative stress
Genotypes for the TNF −308 and LTA NcoI polymorphisms were successfully determined for 212 and 210 active coal miners respectively, and for the 19 retired miners with CWP. As expected, strong linkage disequilibrium was observed between TNF −308 and LTA NcoI genotypes (data not shown). No difference in genotype distributions was observed according to the geographical origin of the miners (France, other European countries, and North Africa). Genotype distributions for the TNF −308 and LTA NcoI polymorphisms fit predictions for Hardy-Weinberg equilibrium.
The mean age of active miners was 47 years and 55 years for retired miners. Half of the active miners were current smokers, 28% were currently exposed to coal dust, and 4% had CWP in 1994. Thirteen percent (13%) of those re-examined five years later (1999) had CWP. In the small group of retired miners, none was currently exposed, approximately half were ex-smokers, and all had CWP in 1994. Age, body mass index, smoking, and current exposure to dust were unrelated to TNF −308 or LTA NcoI genotypes (table 1). Cumulative dust exposure was also unrelated to the TNF –308 genotype. A relation of borderline significance between homozygotes or heterozygotes for LTA NcoI allele 1 with cumulative exposure was observed, which remained after adjustment for age (p=0.07).
TNF −308 and LTA NcoI polymorphisms and environmental exposure to oxidative stress in coal miners
Stages of disease
TNF −308 genotypes were unrelated to 1994 x ray grade or to the CT scan micronodule score with 1999 x ray grade (table 2). In contrast, CT scan micronodule score significantly varied according to LTA NcoI genotypes, even though the LTA NcoI genotype was unrelated to 1994 x ray grade (table 2). A statistically significant relation of LTA NcoI genotypes with 1999 x ray grade was found. In 1999, homozygotes for the LTA NcoI allele 1 had the lowest prevalence of normal x ray grade (35.3%) and subjects with x ray grade 2/1 or over were only found in homozygotes for allele 2 both in 1994 and in 1999. The single subject with 3/+ x ray grade was heterozygous for TNF −308 and LTA NcoI.
TNF −308 and LTA NcoI polymorphisms and stages of pneumoconiosis in coal miners
Markers of response to oxidative stress
Overall, subjects homozygous or heterozygous for TNF −308 allele 2 had significantly lower GSH-Px activity and higher erythrocyte catalase activity (the association was statistically significant after exclusion of two outliers) (table 3). No association was found between biological markers and LTA NcoI genotypes.
TNF −308 and LTA NcoI polymorphisms and biological markers of response to oxidative stress in coal miners in 1994
Gene-environment interaction on GSH-Px activity
We previously found, with the same study sample, a borderline significant association of erythrocyte GSH-Px activity and a significant association of catalase activity with occupational exposure to coal dust.21 In the present study, erythrocyte GSH-Px activity was significantly lower in homozygotes or heterozygotes for TNF −308 allele 2, and we hypothesised that environmental exposure to oxidants (cigarette smoke, dust exposure) interacts with the TNF –308 genotype to determine the capacity of GSH-Px to respond to oxidative stress. Therefore, we studied the association of erythrocyte GSH-Px activity with smoking habit and occupational exposure to dust according to TNF –308 genotypes.
No significant interaction was found between TNF −308 genotypes and smoking habits on erythrocyte GSH-Px activity. Nevertheless, among the miners homozygous or heterozygous for TNF –308 allele 2, mean (SD) erythrocyte GSH-Px activity was negatively and significantly related to smoking habits (44.2 (10.8) U/g Hb in non-smokers, 36.7 (12.6) U/g Hb in ex-smokers, and 34.7 (11.4) U/g Hb in smokers, p=0.05, trend test). Among the miners homozygous for TNF −308 allele 1, mean GSH-Px activity was unrelated to smoking habits (43.3 (12.7) U/g Hb in non-smokers, 39.1 (15.6) U/g Hb in ex-smokers, and 42.5 (12.6) U/g Hb in smokers, p=0.27). Among miners genotyped for TNF –308, a borderline significant association was found between smoking habits and mean estimated cumulative exposure to dust (58.9 (46.0) mg/m3 × year in non-smokers, 66.5 (45.8) mg/m3 × year in ex-smokers, and 51.1 (35.9) mg/m3 × year in smokers, p=0.06).
A significant interaction between cumulative dust exposure and TNF −308 genotypes was found (fig 1). In miners with low cumulative dust exposure, no significant difference was found in erythrocyte GSH-Px activity between miners homozygous or heterozygous for TNF −308 allele 2 and those homozygous for TNF −308 allele 1. In contrast, erythrocyte GSH-Px activity was significantly lower in miners homozygous or heterozygous for TNF −308 allele 2 and with high cumulative dust exposure. A multivariate linear regression model of GSH-Px activity (dependent variable) on cumulative exposure (low v high) and TNF −308 (11 v 12 or 22) was used to test statistically the interaction (that is, the heterogeneity of the association of GSH-Px activity with TNF −308) of exposure × TNF –308, and found a p value of 0.06 (0.05 in a model also including smoking) for the interaction. In those with high cumulative dust exposure, differences according to TNF –308 genotypes were similar when considering separately those with and without CWP in 1994. In miners without CWP, mean (SEM) GSH-Px activity was significantly lower in miners homozygous or heterozygous for TNF −308 allele 2 (36.2 (2.8), n=17) compared with those homozygous for allele 1 (44.6 (2.0), n=43, p=0.02). In the small group of miners with CWP, mean GSH-Px activity was also lower in miners homozygous or heterozygous for TNF −308 allele 2 (23.4 (5.7), n=3) versus those homozygous for allele 2 (52.1 (11.4), n=3, p=0.09). Five year CWP incidence between TNF –308 genotypes in miners with low cumulative dust exposure was similar in miners homozygous for TNF – 308 allele 1 and in those homozygous or heterozygous for TNF −308 allele 2, whereas in those with high cumulative dust exposure, more than two-fold (although not statistically significant) higher CWP incidence was observed. No interaction was found between LTA NcoI genotypes with environmental exposure to oxidants on erythrocyte GSH-Px or catalase activities.
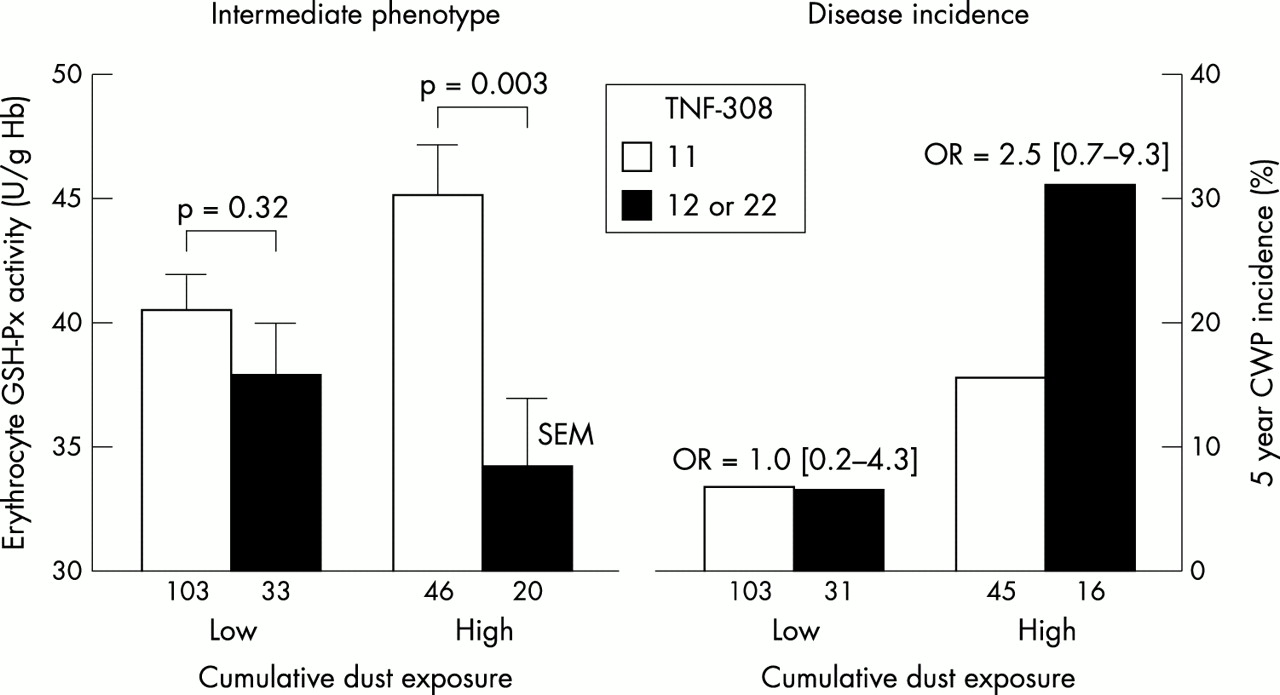
Associations of TNF –308 genotypes with quantitative phenotype of oxidative stress and pneumoconiosis incidence according to cumulative occupational exposure. Cumulative coal dust exposure: low = ≤71 mg.m3 × year, high = >71 mg.m3 × year based on job history and ambient measurements in the coal mine. Numbers of miners (all active in 1994) in each group are shown below each bar. Five year incidence refers to the proportion of those with CWP in 1999 among those without in 1994. OR: odds ratio (95% confidence interval).
Gene intermediate phenotype interaction on prevalence of CWP
We also previously showed21 that erythrocyte catalase activity was positively and significantly associated with 1994 x ray grade. We therefore hypothesised that an interaction exists between LTA NcoI genotypes and erythrocyte catalase activity on disease phenotypes. To test this hypothesis, we studied the association of LTA NcoI genotypes with CT scan score or CWP prevalence according to erythrocyte catalase activity. Miners were classified according to low or high catalase activity, with a threshold based on the third tertile. Owing to the small number of miners with CWP homozygous for the LTA NcoI allele 1, miners homozygous or heterozygous for the LTA NcoI allele 1 were combined and compared with miners homozygous for LTA NcoI allele 2.
Significant interactions between catalase activity and LTA NcoI genotypes on CWP prevalence in 1994 and 1999 were found (fig 2). In miners with low catalase activity, the CWP prevalence was significantly higher in miners homozygous for the LTA NcoI allele 2, whereas in miners with high catalase activity, the CWP prevalence was lower in miners homozygous for the LTA NcoI allele 2. At both surveys, odds ratios were greater than 1 in those with low catalase activity, that is, LTA NcoI 22 genotype increased the risk of pneumoconiosis, but odds ratios were lower than 1 in those with high catalase activity. No interaction was found between LTA NcoI genotypes with catalase activity on CT scan score, or between TNF –308 genotypes with catalase activity on disease phenotypes.

{kind=link}
{kind=link}
Associations of LTA NcoI genotypes with pneumoconiosis prevalence according to the level of quantitative phenotype of oxidative stress. Erythrocyte catalase activity in 1994. Low ≤ 107 k/g Hb, high: = 107 k/g Hb (highest tertile). Numbers of miners (either active or retired in 1994) in each group are shown below each bar. OR: odds ratio (95% confidence interval). Breslow and Day test for interaction refers to the heterogeneity of odds ratios according to catalase activity.
DISCUSSION
In this genetic epidemiological follow up study, we tested the general hypothesis that TNF –308 and LTA NcoI polymorphisms modify pulmonary responses to oxidants in coal miners differently exposed to cigarette smoke and coal mine dust. Results indicated significant gene-environment interaction on intermediate phenotypes (response to oxidative stimuli), and gene-intermediate phenotype interaction on clinical disease (coal worker’s pneumoconiosis). An interaction of the TNF −308 genotype with quantitatively assessed occupational exposure on erythrocyte GSH-Px activity was observed, with a significant association in those with high exposure, whereas no association was found among those with low exposure. Further, assessment of the interaction of gene and biological response to oxidative stress on clinical phenotype identified a significant association of CWP prevalence with LTA NcoI in those with low catalase activity whereas no association was observed in those with high (a priori protective) activity.
Assessing gene-environment interaction has proven to be difficult, and few clear examples have been reported and replicated. Problems often encountered are the lack of specific hypotheses (candidate interactions), insufficient characterisation of exposure, and the availability of sufficiently low level (closer to the gene than clinical disease) phenotype markers. Unravelling biochemical and physiological hierarchies leading from genes to clinical end points in complex diseases need new strategies.30 This study attempted to incorporate a conceptual temporal sequence31 from environment and genes towards disease. In such an overall framework of aetiological factors to severe disease, it is possible to distinguish (1) internal dose (inhaled coal dust), (2) early biological effect (low level intermediate phenotype30 such as enzymatic response to oxidative stimuli), (3) altered structure/function (CT score), and (4) clinical (x ray confirmed CWP) and severe (nodular form) disease. We attempted to apply this framework to study gene-environment interaction and gene-intermediate phenotype interaction in the pathogenesis of CWP.
Strengths of the study were the contrasted exposure by design in the study sample, the availability of quantitative objective measures of the coal mine dust, the availability of various quantitative phenotypes to assess response to oxidative stress, subclinical computed tomography score, and two assessments of CWP by standardised x ray five years apart. Limitations of the study were the relatively small sample size that precludes detailed analyses addressing simultaneously the various steps of exposure, as well as intermediate and clinical phenotypes. Another limitation is the particular type of the population studied (coal miners), but it does permit study of a homogeneous group of subjects. The window of opportunity for evaluation of the role of single or multiple genes in disease pathology may depend on the relative timing of the environmental stimuli, but also the stage of the disease. Furthermore, genetic background may play a key role in the occurrence and/or the severity of the disease.32,33 In the present study, it was not possible to address the role of TNF and LTA in the pathogenesis from established CWP towards the most severe forms, but continued follow up of the study sample will allow us to address this question later.
Another important consideration in a study of gene-environment interaction and disease pathogenesis is the complex quantitative nature of the disease under study. Almost certainly, multiple genes contribute to CWP susceptibility. However, owing to the small size of the study population, the investigation was limited to those genes with relatively common polymorphisms with known roles in chronic pulmonary diseases (see below). That is, the study population does not have sufficient statistical strength to evaluate potentially important rare gene polymorphisms. The TNF –308 and LTA NcoI polymorphisms are relatively common in European populations (approximately 14% and 30%, respectively, in the present study), and each has demonstrated roles in chronic pulmonary diseases and responses to oxidant stimuli. For example, the TNF –308 polymorphism has been associated with chronic bronchitis,10 fibrosing alveolitis,11 asthma,34 and a higher risk of poor prognosis in COPD.35 TNF-α has also been shown to be an important determinant of susceptibility to lung inflammation and injury induced by the environmental oxidant ozone (O3) in animal models.36 Similarly, Shimura et al37 reported that LTA NcoI allele 2 was less prevalent in 135 lung cancer patients as compared to 165 normal healthy subjects. The authors also reported that among patients with stage III and IV lung cancer (n=31), those homozygous for the LTA NcoI allele 1 (n=9) had a two fold greater survival rate five years after operation than in patients homozygous or heterozygous for the LTA NcoI allele 2, whereas no difference was found among patients with stages I and II. Campbell et al38 found a significant protection of subjects homozygous for LTA NcoI allele 1 against recurrent wheeze and a borderline protection against asthma in a random cohort of 224 subjects. Further, Stuber et al28 reported a significant association of LTA NcoI allele 2 with worse outcome among 40 patients with severe sepsis. However, Yamaguchi et al18 found that LTA NcoI allele 1 is associated with a prolonged clinical course of sarcoidosis in 110 Japanese patients and other studies reported no association with disease.34,39 Ethnic differences or the fact that some of these patients showed mild to moderate disease manifestations may partly explain these discrepancies.
Several arguments support the hypothesis that markers of the response to oxidative stress are relevant intermediate phenotypes for CWP. It is well known that CWP is related to increased amounts of hydrogen peroxide40,41 and catalase converts hydrogen peroxide to water and oxygen. We previously found that catalase activity was related to pneumoconiosis and was higher in miners with a CT micronodule score ≥ 4.21 In a five year follow up study conducted in 66 coal miners, incidence or disease progression was associated with the combination of initial levels of various antioxidant enzyme activities.42 Similarly, a potential role of TNF genotype on GSH-Px response to oxidative stimuli is consistent with previous studies. TNF-α is known to generate ROS by leakage from the mitochondrial electron transport chain43 and modulates glutathione levels (GSH), one key component for GSH-Px activity. Moreover, the gene for glutathione peroxidase is induced in response to TNF-α in lung cells.44 This may have important implications for the oxidant/antioxidant balance in inflammatory lung diseases. Overall, because of the systemic nature of the response to oxidative stress studied, the framework applied in this study is of general interest for other diseases that depend on the same pathways.
The present study showed an interaction of the TNF −308 genotype with high coal dust exposure on GSH-Px activity. As the less common allele 2 is associated with higher basal and induced expression of TNF-α,45 and TNF-α and ROS cooperate to activate transcription factors,46 we may postulate that TNF-α modulates GSH-Px activity through the regulation of GSH levels. In the presence of chronic oxidative stress resulting from high cumulative dust exposure, antioxidant defences may be overwhelmed, leading to alteration of the antioxidant enzymes as lower erythrocyte Cu++/Zn++ SOD activity, as we observed previously,21 and down regulation of GSH-Px.47 Erythrocyte GSH-Px activity plays a major role in ROS metabolism in most tissues and has been shown to control NF-κB transcription factor activation.48 As plasma GSH-Px originates from the kidney,49 it mostly plays a role in response to acute oxidative stress, which is consistent with our observation that only erythrocyte GSH-Px response depends on cumulative exposure.
Our results support and extend findings from a Belgian study in which TNF −308 allele 2 was more prevalent in 18 ex-coal miners with x ray grade 0/1 or over (14 of whom had x ray 1/1 or over) as compared to 60 ex-coal miners with normal x ray (0/0) and 56 controls.13 Previously, they reported that spontaneous and coal dust stimulated TNF release from peripheral blood monocytes was increased in coal miners with early CWP.50 Furthermore, they found that miners who showed abnormally high release of TNF stimulated by coal dust had an increased risk of progression in CWP over five years.51 Similarly, TNF −308 allele 2 was found to be more prevalent in 80 patients who had CWP than in 54 healthy controls, with a twice higher prevalence in those with large opacities as compared to those with simple CWP.14 A necropsy study, in which the severity of fibrosis was anatomically assessed, showed a significant prevalence of allele 2 restricted to moderate fibrosis in 325 former coal miners as compared to 164 controls.52 In contrast, the association of allele 2 with disease was restricted to severe silicosis (nine cases) in 241 South African gold miners.33 In none of these studies was the analysis stratified according to dust exposure to assess gene-environment interaction. Limited information is available on the association of TNF with the first stages of the disease. No association in the present study between TNF –308 genotypes and CT score was found, but it is worth noting that although it was not statistically significant, a two-fold increase in five year CWP incidence was observed in miners homozygous or heterozygous for TNF −308 allele 2 with high cumulative dust exposure.
Overall, our findings and previous results on the severity of various diseases (in particular, silicosis) implicate TNF –308 at various stages of disease progression, but further work is needed to clarify mechanisms and interaction with other genes. Another important consideration is that CWP, similar to other complex diseases, probably involves numerous genes acting at each step. While a good rationale is provided for the study of TNF and LTA, other genes implicated in the development of oxidative stress, such as antioxidant enzyme genes and transcription factors that modulate their expression, should be investigated. Further, well designed studies that take advantage of emerging expression array and proteomics technologies in human subjects and animal models exposed to coal dust, or oxidant mixtures, should yield insight into other important candidate susceptibility genes.
In conclusion, this is the first demonstration of the involvement of functional polymorphisms of two candidate genes TNF and LTA in the control of the physiological/biochemical responses to quantitatively assessed oxidative stimuli. The study design enabled investigation of environment and disease interactions, and provides a better understanding of the aetiology of the chronic inflammatory lung diseases such as pneumoconiosis.
Acknowledgments
The paper is dedicated to the late Quang-Thieu Pham who set up the coal miner study. We thank the medical staff and all the French coal mine workers who made this study possible. We also thank A Berthelin, M Dusch, and G Hellier for their skilful technical assistance. This research was in part supported by grant numbers 7280.03.001 and 7280.03.035 from the European Community of Steel and Coal, National Institutes of Health grant ES-09606, and Environmental Protection Agency grant R-825815.
REFERENCES
Footnotes
-
↵* Present address: Laboratory of Pulmonary Pathobiology, National Institute of Environmental Health Sciences, Research Triangle Park, NC, USA